|
NOVAS
ALTERNATIVAS PARA GERMOPLASMA FORRAGEIRO - LEGUMINOSAS
Paulo
Bardauil Alcântara 1. INTRODUÇĂO No Brasil, na década de 70, a introduçăo, avaliaçăo e seleçăo de germoplasmas baseavam-se no processo de coleta de espécies nativas e exóticas presentes na natureza. Porém, com o decorrer dos anos tornou-se um processo lento e de alto custo para a sociedade científica. Atualmente, um novo conceito sobre melhoramento genético de plantas forrageiras vem sendo consolidado – o processo de desenvolvimento de cultivares. Esse processo é uma das atividades mais relevantes da pesquisa agropecuária brasileira, apresentando grandes retornos sociais e econômicos para os clientes dos segmentos público e privado atuantes no setor (CASTRO et al, 2002). Năo podemos prever as surpresas que os próximos anos nos trarăo, inclusive pela inesperada rapidez com que ocorrem, mas a aprovaçăo das novas legislaçőes de proteçăo ŕ propriedade intelectual (lei de patentes e proteçăo de cultivares) e os avanços nas técnicas de melhoramento genético utilizando tecnologia de ponta permitirăo uma maior eficięncia na produçăo animal. Recentemente Brasília sediou o Workshop Internacional sobre Desenvolvimento da Agricultura Tropical. Neste evento levantou-se a questăo do esquentamento global provocado pelo efeito estufa. De fato houve um aumento médio de 1şC na temperatura da Terra o que levou ao avanço de 110 km para o norte e para o sul do equador aumentando a área tropical do planeta. Com isso tem-se também uma extensăo maior de desertos. O prognóstico é que os trópicos, cada vez mais presentes na geografia do globo, serăo os grandes responsáveis pela produçăo agrícola para o mundo. Dessa maneira urge um programa que contemple novas tecnologias visando o aumento significativo da produtividade nas várias cadeias do agronegócio. Neste aspecto segundo REIS VELLOSO (2006) o Brasil possui um trunfo único no mundo que é o de ampliar a fronteira agrícola sem se utilizar do desmatamento e assim sem avançar sobre a Amazônia. Cerca de 90 milhőes de hectares podem ainda ser incorporados ŕ área produtiva, o que corresponde a 1,5 vezes o que já temos sob cultivo. Segundo o mesmo autor, há que se vencer tręs obstáculos: a logística, a precariedade dos corredores de exportaçăo e os constantes cortes na área de desenvolvimento tecnológico. Muito embora a adoçăo de leguminosas em pastagens ainda seja tímida, testemunhamos atualmente um crescente interesse da iniciativa privada pelas leguminosas. Tal interesse iniciou-se nas empresas sementeiras principalmente naquelas com maior visăo de futuro e maior conhecimento técnico e vem se alastrando como uma onda positiva dentre os pecuaristas que procuram um produto diferenciado através de pastagens de melhor qualidade. Várias săo as causas apontadas para a baixa adoçăo das leguminosas: a) falta de percepçăo dos benefícios causados por essas plantas; b) falhas de tecnologia (cultivares inadequados, manejo errado, falha no plantio e estabelecimento, etc.); c) fatores sócio-econômicos (tecnologia requerida é mais cara, tradiçăo, regime pluviométrico muito variável, etc.); d) falhas no enfoque (lançamento errados, falta de sementes, falta de parcerias efetivas entre o público e o privado, falta de um programa sólido para produçăo de sementes, preços irreais). Paralelamente, a essas razőes que motivam a baixa adoçăo de leguminosas, há outras que inauguram uma história futura de sucesso nos anos vindouros. As dificuldades para promover o uso de leguminosas e conseqüentemente as baixa taxas de adoçăo das mesmas como plantas forrageiras tem sido a grande preocupaçăo das instituiçőes de P, D&I. É necessário aumentar o interesse por essas espécies demonstrando de maneira explícita seus benefícios para que năo retrocedam os interesses dos órgăos financiadores de pesquisa e da iniciativa privada. (SHELTON et al., 2005). Esses autores apontam como fatores de sucesso no uso de leguminosas os seguintes: a) a tecnologia atende ŕs necessidades dos pecuaristas embora elas variem de um para outro; b) tecnologia adaptada ŕs situaçőes sócio-econômicas e ao nível do produtor; c) parcerias efetivas e de longo prazo entre órgăos oficiais e a iniciativa privada (SHELTON et al, 2005). Independentemente das açőes sempre positivas das instituiçőes de pesquisa, é necessário agregar-se ao programa um intenso trabalho de difusăo e transferęncia dessas novas tecnologias para que a adoçăo das leguminosas saia da timidez exagerada e alcance seu devido lugar na produçăo da carne verde. Săo cada vez mais raros os lançamentos de cultivares de forrageiras oriundos de ecótipos coletados na natureza. Isto decorre de inúmeras causas dentre as quais podem ser listadas o alto custo das expediçőes de coleta, a falta de intercâmbio de material genético entre os órgăos de pesquisa devido ŕ lei de proteçăo de cultivares, a erosăo genética, a destruiçăo dos ecossistemas e dos nichos ecológicos, a contaminaçăo genética e o eminente esgotamento dos acessos existentes nos bancos de germoplasma. Há ainda muitos acessos depositados em bancos de germoplasma sem a adequada avaliaçăo principalmente pela falta de recursos ou de prioridades. Com a crescente demanda pela busca de plantas mais produtivas, tolerantes a estresses biológicos, de alto valor nutritivo, com aceitabilidade, de alta persistęncia, etc. é de se esperar que programas permanentes de melhoramento e avaliaçăo sejam implantados no país. Várias metodologias vem sendo aplicadas ŕs leguminosas. Algumas delas já tradicionalmente usadas, outras bem mais recentes surgem como possível soluçăo para os problemas de maneira mais rápida. 1) Avaliaçăo de ecótipos; 2) Melhoramento clássico; 3) Melhoramento assistido por marcadores moleculares e 4) Transgenia.
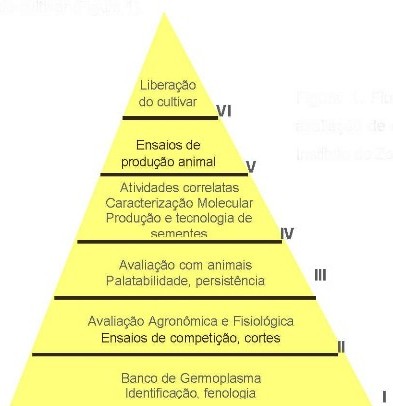
As etapas principais de um programa de seleçăo e melhoramento de forrageiras utilizadas pelo Instituto de Zootecnia săo: I) Banco de Germoplasma (Identificaçăo, fenologia); II) Avaliaçăo agronômica e fisiológica (Ensaio de competiçăo, cortes); III) avaliaçăo de grandes parcelas sob pastejo (Palatabilidade, persistęncia); IV) Atividades correlatas, caracterizaçăo molecular e produçăo e tecnologia de sementes; V) Ensaio com desempenho animal; VI) Liberaçăo do cultivar (Figura 1)
Segundo DALL’AGNOL & SCHEFF-BASSO (2004) tais fases săo conceitualmente comuns na maioria dos programas de melhoramento genético nos trópicos, porém algumas das etapas tęm sido omitidas. Com a intençăo de se expor os materiais o mais rapidamente possível para os produtores, muitas avaliaçőes vęm sendo realizadas ao nível de fazendas. Para esses autores, algumas consideraçőes devem ser feitas acerca da metodologia de avaliaçăo do germoplasma de leguminosas tropicais, no sentido de aperfeiçoar tal processo, principalmente ao nível da caracterizaçăo e avaliaçăo preliminar. Algumas sugestőes incluem: a) identificaçăo precoce de duplicatas genéticas, b) determinaçăo do sistema de fecundaçăo e otimizaçăo do manejo de populaçőes alógamas, c) determinaçăo da herdabilidade para os principais descritores, d) diagnóstico ilustrado das principais doenças, e) determinaçăo rápida dos efeitos da melhoria do solo sobre as plantas. A utilizaçăo de métodos clássicos como a introduçăo, seleçăo fenotípica e seleçăo genotípica ainda contribuirăo bastante para os avanços no melhoramento genético, quando agregadas com as novas técnicas biotecnológicas. Assim, o sucesso futuro do melhoramento dependerá da utilizaçăo integrada das técnicas biotecnológicas com métodos convencionais de melhoramento, necessitando, dessa forma, de uma estreita colaboraçăo entre geneticista molecular e melhorista (PEREIRA, 2002).
O processo de hibridaçăo é uma das etapas mais importantes no melhoramento genético de uma espécie vegetal. A hibridaçăo é essencial para o desenvolvimento de novas variedades, uma vez que de cruzamentos de parentais geneticamente distintos săo desenvolvidas populaçőes com variabilidade genética, para aplicaçăo de métodos apropriados de avaliaçăo e seleçăo de características superiores. Com objetivos preestabelecidos, săo realizadas hibridaçőes para criar germoplasma diverso e fazer frente aos problemas reais ou potenciais da cultura. Um cultivar altamente produtivo e estável representa uma combinaçăo bem balanceada de genes, interagindo com o ambiente em que é cultivado. As hibridaçőes săo feitas na forma de cruzamentos simples, duplos ou múltiplos e servem para recombinar a variabilidade genética intra-específica ou para aumentar essa variabilidade, pela introgressăo de genes em cruzamentos interespecíficos (PINTO, 2001).
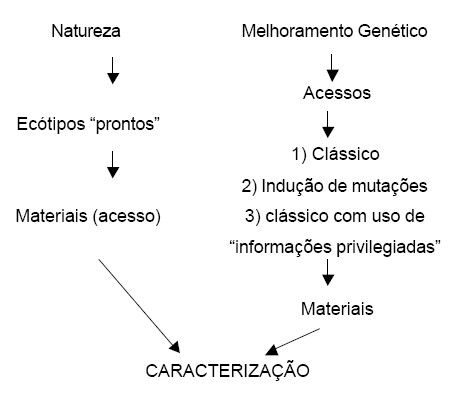
Os métodos clássicos como a introduçăo, seleçăo fenotípica e seleçăo genotípica ainda săo bastante utilizados nos avanços do melhoramento genético, principalmente, quando agregadas com novas técnicas (Figura 2).
Método Clássico assistido por marcadores moleculares As etapas fundamentais de um programa de melhoramento de plantas săo: obtençăo da variabilidade genética; seleçăo de indivíduos superiores; e avaliaçăo de materiais genéticos promissores para lançamento comercial. Apesar de ganhos genéticos significativos terem sido obtidos na seleçăo de características de interesse na maioria das espécies de importância agronômica, a expectativa de progresso genético e obtençăo de indivíduos ainda melhores permanecem. Por esta razăo, os melhoristas de plantas tęm buscado formas de continuar obtendo ganhos genéticos, tornando mais eficiente cada uma das etapas do programa. A essęncia deste progresso está em o quanto o fenótipo de um indivíduo expressa o seu genótipo. Por isso que o uso de marcadores moleculares pode auxiliar o melhoramento de plantas, pois possibilita acessar diretamente o genótipo de um indivíduo. Assim, entre as principais aplicaçőes de marcadores de DNA em programas de melhoramento de plantas estăo: o monitoramento e organizaçăo da variabilidade genética e a seleçăo assistida por marcadores moleculares. Os marcadores moleculares terăo grande impacto e utilizaçăo em algumas situaçőes, mas năo em todas. Assim, cabe aos melhoristas junto com os especialistas na área molecular decidirem caso a caso se devem ou năo utilizar esta tecnologia. O difícil e nem sempre claro é responder “quando, em que situaçőes, utilizar marcadores moleculares”. Em termos de variabilidade genética, marcadores moleculares permitem: compreender e organizar a variabilidade genética de um programa de melhoramento de forma única, isto é, acessando variabilidade de DNA, que năo é influenciado pelo ambiente como săo, por exemplo, os caracteres morfológicos e fenotípicos em geral de uma planta. A primeira conseqüęncia disto é a possibilidade de planejar os cruzamentos de um programa de forma a maximizar as diferenças genéticas entre genótipos elites, diferenças essas que muitas vezes năo podem ser observadas em nível de fenótipo. A segunda é a possibilidade de organizar o germoplasma do programa em pools gęnicos (grupos de genótipos com características comuns dentro de uma espécie), facilitando a escolha e diminuindo o número de combinaçőes a serem feitas pelo melhorista (YAMAME, 2002). O que melhorar? Os programa de melhoramento de leguminosas forrageiras tropicais deve ser direcionado para a seleçăo de novos materiais que possam aumentar a qualidade e a produtividade da forragem produzida e a eficięncia da produçăo animal levando-se em consideraçăo o ambiente. Os gęneros de leguminosas arbóreas e arbustivas que vęm sendo estudados no Brasil săo Cajanus, Leucaena, Sesbania e a Cratylia. Quanto aos gęneros herbáceos podem ser citados o Arachis, Calopogonium, Centrosema, Macroptilium, Galactia, Stylosanthes e a Neonotonia. As características de maior interesse no processo de seleçăo das leguminosas săo: qualidade, produçăo de sementes, resistęncia a pragas e doenças, fixaçăo de nitrogęnio, persistęncia e tolerância á seca e ao frio. A busca por melhorias na qualidade das leguminosas forrageiras tem sido considerada como um dos objetivos do melhoramento genético. No caso das leguminosas arbóreas e arbustivas pesquisas tem sido conduzidas para seleçăo de espécies de leguminosas que possuam menos fatores antiqualitativos e praticamente livres de substâncias estrogęnicas. Por outro lado, quando o assunto é a consorciaçăo entre gramíneas e leguminosas herbáceas estudos vem sendo conduzidos para a seleçăo de espécies de leguminosas que tenham aceitabilidade menor em relaçăo ŕ gramínea associada, visando sua maior persistęncia durante a estaçăo chuvosa e, por outro lado, sendo bem aceita pelos animais na época crítica (período de escassez de forragem). Outra característica das leguminosas que concorre para assegurar sua persistęncia em pastagens é a produçăo adequada de sementes viáveis, de modo a manter uma reserva das mesmas no solo. A germinaçăo de uma proporçăo dessas sementes a cada estaçăo de crescimento concorre para manter a populaçăo de plantas em nível desejável e a estabilidade da pastagem, sendo importante mencionar a viabilidade econômica de sua implantaçăo, no sentido de baratear o custo.
Alguns gęneros de
leguminosas como o Cajanus
vem sendo alvo de estudo de melhoramento genético pelo Instituto de
Zootecnia em parceria com o Instituto Agronômico de Campinas no sentido
de obter cultivares com porte menor, arquitetura mais ereta e com ramos
que se verguem e năo quebrem, com boa produçăo de massa foliar e,
principalmente de sementes. E também está sendo dada preferęncia ŕ
seleçăo por plantas mais precoces ou mais tardias quanto ŕ produçăo de
sementes. 4. TRANSGENIA As técnicas de transformaçăo genética podem ser consideradas a continuaçăo de uma longa lista de métodos tradicionais de melhoramento, como a induçăo de mutaçőes, a hibridizaçăo entre espécies e gęneros, a duplicaçăo de cromossomos, a cultura de células e tecidos in vitro e a fusăo de células somáticas. A modificaçăo de bactérias, animais e plantas com essas novas técnicas, superando as barreiras naturais, vem gerando muita polęmica. Entretanto, por sua novidade, a transgenia tem causado preocupaçőes e transformaçőes, sejam de caráter científico, social, econômico ou cultural. Assim, a controvérsia provocada pelas novas técnicas da biotecnologia, em especial o uso de organismos transgęnicos ou de organismos geneticamente modificados (OGMs), deve ser considerada uma mudança na percepçăo pública sobre a cięncia e sobre as conseqüęncias das suas aplicaçőes tecnológicas (http://cienciahoje.uol.com.br). Atualmente técnicas de transgenia săo capazes de eliminar, inserir e transferir genes permitindo a construçăo de diversos tipos de organismos com o genoma modificado, como, por exemplo, bactérias e até animais capazes de produzir certas proteínas de interesse para a saúde humana (como a insulina e hormônios de crescimento), camundongos e outros animais de laboratório sem determinados genes ou com genes alterados (destinados a pesquisas de vários tipos), e uma ampla variedade de plantas com características especiais, como resistęncia ao ataque de pragas e doenças, tolerância a diversos estresses ambientais (seca, frio, salinidade etc.), maiores teores de aminoácidos essenciais, de vitaminas e de compostos com açăo farmacológica, além de alteraçőes na coloraçăo, no sabor ou em aspectos físicos e químicos dos alimentos (VIEIRA, 2004). Apesar das grandes discussőes em torno dos OGMs, as plantas geneticamente modificadas representam hoje um caminho promissor para o melhoramento vegetal. Inúmeros exemplos de estratégias de transferęncia de genes conferiram, com sucesso, resistęncia a herbicidas, vírus, fungos, bactérias e insetos ou produziram um aumento na qualidade dos alimentos. Além das aplicaçőes biotecnológicas, as plantas transgęnicas tęm contribuído significativamente para o estudo do funcionamento dos genes, tais como a análise da regulaçăo da expressăo gęnica e o estudo das funçőes das proteínas codificadas pelos diferentes genes da planta (PINHEIRO et al., 2000). Independentemente do método de transformaçăo utilizado, a primeira etapa para se obter a planta modificada requer a disponibilidade do gene que codifica a característica a ser introduzida no vegetal, assim como a porçăo reguladora do gene ou promotor, o qual irá determinar os tecidos e as fases do desenvolvimento nos quais o gene será expresso. Dessa maneira, é possível construir genes quiméricos, que contęm promotores diferentes daqueles dos genes originais. Na maioria das plantas transgęnicas já obtidas e comercializadas, a característica de interesse é expressa em todos os tecidos, năo importa a fase de desenvolvimento em que se encontra o vegetal, graças ŕ utilizaçăo de um promotor do vírus do mosaico da couve-flor, o 35S do CaMV. Além do gene de interesse sob o controle de um promotor, o fragmento a ser transferido (construçăo ou cassete para transformaçăo) deve conter também um gene de seleçăo ou marcador. Esse gene irá conferir ŕ planta resistęncia a um antibiótico ou herbicida, de maneira que somente aquelas que efetivamente receberam o cassete de transformaçăo crescerăo em um meio que contém os agentes seletivos específicos. Grande número de plantas transgęnicas tęm sido gerado e submetido a testes de campo em muitos países. Todas essas plantas contęm um gene marcador, que tem sido fonte de muitos questionamentos sobre a segurança — para o ambiente e para o consumidor — das plantas geneticamente modificadas. Recentemente, foram descritas algumas técnicas que visam a retirar o gene marcador, como, por exemplo, a utilizaçăo de vetores contendo dois T-DNAs separados para a co-transformaçăo via A. tumefaciens e segregaçăo dos transformantes livres dos marcadores (KOMARIT et al., 1996). No entanto, essas técnicas ainda năo foram utilizadas em aplicaçőes comerciais. É preciso ressaltar que elas năo podem ser aplicadas para as lenhosas, para as plantas propagadas vegetativamente ou para aquelas estéreis, pois o cruzamento sexual é essencial para segregar o gene marcador seletivo e o gene de interesse. Sob condiçőes especiais de cultivo, cada célula transformada poderá ser regenerada em uma planta completa, dando origem a uma linhagem transgęnica. Cada linhagem conterá uma ou mais inserçőes em seu genoma. Esse processo de integraçăo é aleatório, ou seja, cada linhagem terá o transgene integrado em diferentes loci. Isto poderá resultar em uma diversidade no nível de expressăo gęnica entre as diferentes linhagens. Por isso, diversas análises genéticas, bioquímicas e moleculares devem ser realizadas, antes que uma planta seja liberada para testes em casas de vegetaçăo ou em campo. Essas análises visam confirmar a integraçăo e a expressăo do gene, além de selecionar as plantas que năo apresentam nenhum tipo de alteraçăo além daquela desejada (SANTARÉM, 2003). Métodos Diretos: Os métodos de transferęncia direta de genes utilizam processos físicos ou químicos que causam modificaçőes nas paredes e membranas celulares, facilitando a introduçăo de DNA exógeno. Eletroporaçăo de Protoplastos Protoplastos săo definidos como células desprovidas de paredes celulares (EVANS, 1991). Para a introduçăo de DNA usando a eletroporaçăo, os protoplastos săo expostos a pulsos curtos de corrente contínua e alta voltagem, em presença do DNA exógeno. Esse tratamento induz uma alteraçăo reversível da permeabilidade da membrana plasmática e poros temporários săo formados, permitindo a entrada do DNA nas células. Bombardeamento de Partículas Esse método consiste na aceleraçăo de micropartículas de metal (principalmente ouro coloidal), que atravessam a parede celular e a membrana plasmática, carreando DNA para o interior da célula. O termo bombardeamento de partículas pode ser substituído por aceleraçăo de microprojéteis ou método biobalístico. O método baseia-se no uso de um equipamento que produz uma força propulsora, usando pólvora, gás ou eletricidade, para acelerar micropartículas inertes, cobertas com DNA, em direçăo ŕs células alvo. Após o bombardeamento, uma proporçăo de células atingidas permanece viável; o DNA é integrado no genoma vegetal e incorporado aos processos celulares de transcriçăo e traduçăo, resultando na expressăo estável do gene introduzido (FINER et al., 1996). A maioria dos modelos biobalístico atuais emprega macroprojéteis, usados como veículo para aceleraçăo dos microprojéteis colocados na sua superfície. O uso do processo biobalístico é bastante amplo e, quando comparado com a maioria dos métodos diretos de introduçăo de DNA em plantas, o bombardeamento de partículas apresenta várias vantagens. É uma técnica altamente versátil e de fácil adaptaçăo, podendo ser aplicada a grande variedade de células e tecidos, incluindo suspensőes (FROMM et al., 1990), calos (VASIL et al.,1985), tecidos meristemáticos (MCCABE & MARTINELLI, 1993), embriőes imaturos (SOUTHGATE et al.,1998) e embriőes somáticos (SANTARÉM & FERREIRA, 1997). Essa técnica tem permitido a regeneraçăo de plantas transgęnicas de maneira reproduzível e com menos variabilidade entre experimentos (LUTHRA et al., 1997). MÉTODO INDIRETO Agrobacterium Agrobacterium é uma bactéria de solo, Gram-negativa , aeróbica, pertencente ŕ Família Rhizobiaceae (ZAMBRISKY, 1988). Sua importância para os estudos de transformaçăo de plantas reside na capacidade natural que esses patógenos possuem de introduzir DNA em plantas hospedeiras. Esse DNA é integrado e passa a ser expresso como parte do genoma da planta. Como conseqüęncia dessa expressăo, o padrăo normal de desenvolvimento é alterado: A. tumefaciens causa a formaçăo de tumores, ao passo que a infecçăo por A. rhizogenes resulta na proliferaçăo de raízes (Lipp-Nissinen, 1993). Os métodos baseados na bactéria Agrobacterium tumefaciens săo os mais eficientes para transformar dicotiledôneas (GELVIN, 1998). Essa bactéria é encontrada no solo e tem como mecanismo de infecçăo a transferęncia de um fragmento de DNA do seu genoma para a célula vegetal. Uma vez introduzido no genoma vegetal, esse fragmento de DNA bacteriano (T-DNA ou DNA de transferęncia) passa a dirigir a síntese de hormônios de crescimento. Forma-se um tumor no local de infecçăo. Utilizando-se a tecnologia do DNA recombinante, é possível manipular o T-DNA e substituir os genes produtores de tumor por outro gene de interesse. Dessa maneira, a infecçăo resultará na expressăo da característica de interesse, e o tumor năo será produzido. A inoculaçăo de células ou pequenos fragmentos de tecidos vegetais com a bactéria, contendo o T-DNA modificado, permitirá que essa célula transformada seja regenerada em uma planta completa, graças ao fenômeno de totipotęncia. O desenvolvimento de protocolos de regeneraçăo é, portanto, indispensável para a manipulaçăo genética de plantas. 4.1. Aplicaçőes no Melhoramento de Leguminosas Resistęncia a Herbicidas A resistęncia a herbicidas é uma característica de grande interesse agronômico por causa do uso intensivo desses compostos na agricultura. A produçăo de plantas capazes de tolerar a exposiçăo a herbicidas pode ser obtida pela introduçăo de genes que codificam enzimas capazes de degradar ou detoxificar o herbicida em questăo. Dependendo do modo de açăo do herbicida, outras estratégias podem ser utilizadas. Alguns herbicidas atuam de modo a inativar enzimas vitais do metabolismo vegetal. Nesse caso, é possível produzir plantas que superexpressem a enzima-alvo ou, alternativamente, expressem uma versăo alterada desta, de forma a tornar a planta insensível ao herbicida (FREYSSINET & DEROSE, 1994). A soja Roundup Ready da multinacional Monsanto é o exemplo mais famoso de modificaçăo genética feita para aumentar a resistęncia a um herbicida. Nesse caso, foi utilizada a estratégia que visa a expressăo de um gene modificado, de modo a produzir uma enzima-alvo insensível ao herbicida glifosato, o qual age como inibidor específico da enzima enolpiruvil-shiquimato-3-fosfatase-sintase (EPSP), que participa da síntese de aminoácidos aromáticos, essenciais ŕs plantas. Resistęncia a Pragas e doenças A produçăo de plantas resistentes a pragas e doenças é uma prioridade dos programas de melhoramento. Várias estratégias tęm sido empregadas, tendo em vista obter plantas transgęnicas mais tolerantes a diferentes agentes agressores. Um dos exemplos mais conhecidos săo as plantas transgęnicas que expressam a toxina produzida pelo Bacillus thuringiensis. O B. thuringiensis é uma bactéria gram-positiva que existe no solo, na superfície das plantas e na poeira dos grăos estocados. Durante a esporulaçăo, essa espécie de Bacillus produz cristais paraesporais que consistem de uma ou mais (-endotoxina ou proteína cristal (Cry) de aproximadamente 130kDa. Após a ingestăo dessa toxina pelo inseto, os cristais dissolvem-se no ambiente alcalino do intestino deste e liberam as protoxinas, que săo subseqüentemente processadas pelas enzimas digestivas, produzindo a toxina ativa. Essa toxina se liga a receptores na membrana das células do epitélio digestivo e insere-se nessas membranas, formando poros que resultam na morte das células epiteliais e, eventualmente, do próprio inseto, por lise osmótica coloidal (KNOWLES & DOW, 1993). Já foram obtidas diversas espécies vegetais que expressam a toxina Bt e săo resistentes a insetos, podendo ser citados o fumo, o tomate, o algodăo, a batata e o arroz. Outra estratégia para obter plantas resistentes a insetos consiste na superexpressăo de proteínas inibidoras de enzimas digestivas. A introduçăo do gene do inibidor de tripsina de caupi (Vigna unguiculata) em tabaco promoveu proteçăo contra a praga Heliothis virescens, que ataca o milho e o algodăo. Outro exemplo consiste na produçăo de ervilhas cujas sementes expressam o gene de um inibidor de -amilase. Essa estratégia resultou em sementes resistentes a carunchos, insetos da família Bruchidae (SHADE et al, 1994). Na maioria das vezes, as estratégias para produzir plantas transgęnicas resistentes a fungos baseiam-se em genes vegetais cujos produtos săo capazes de inibir diretamente o crescimento de fungos, como, por exemplo, as quitinases, as glucanases, a osmotina, as lectinas e as tioninas (CORNELISSEN & MELCHERS, 1993). Considerável é o número de culturas transgęnicas produzidas visando desenvolver resistęncia ŕ infecçăo por vírus vegetais (HADIDI et al, 1998). Foram empregadas diferentes estratégias que, na sua maioria, utilizam seqüęncias nucleotídicas e a expressăo de proteínas do próprio vírus. Para o vírus do enrolamento das folhas da batata (PLRV, Potato LeafRoll Virus), dos anéis necróticos do mamoeiro (PRSV, Papaya RingSpot Virus), entre muitos outros, a superexpressăo da proteína da capa do vírus conferiu resistęncia a variedades transgęnicas (GONSALVES & SLIGHTOM, 1993). Tolerância a Alumínio A produçăo agrícola em solos ácidos, que constituem quase a metade do total da superfície arável da Terra, tem como principal limitaçăo a toxidez por Alumínio (Al+3), que se manifesta na reduçăo do crescimento radicular, impedindo a absorçăo de nutrientes e água do solo. Estes solos estăo localizados principalmente no Sudeste Asiático, na América Latina e na regiăo próxima ao Saara na África. Coincidindo com a localizaçăo de áreas em que a utilizaçăo de insumos agrícolas, como a aplicaçăo de calagem para a neutralizaçăo do solo e, conseqüente insolubilizaçăo de Al+3, é restrita. O uso de variedades tolerantes ao Al+3 restaura a capacidade produtiva de regiőes de solos ácidos com altos teores de Al+3. Estudos recentes mostraram que a introduçăo de determinados genes promoveu aumento moderado a elevado na tolerância ao Al+3 em mono e dicotiledôneas susceptíveis. A toxidez por Al+3 é a principal limitaçăo para a produçăo em 37,9% das pastagens do Sudeste da Ásia, 30,9% da América Latina e aproximadamente 20% da Ásia Oriental, da África Sub-Saara e da América do Norte (WOOD et al., 2000). As forragens mais utilizadas nestas regiőes săo gramíneas do gęnero Brachiaria, principalmente Brachiaria brizantha (C. Hochstetter ex A. Rich.) R. Webster, consorciada com leguminosas forrageiras de maior valor nutricional, como soja perene (Neonotonia wightii (Wight & Arn.) Lackey, leucena (Leucaena leucocephala (Lam.) de Wit), estilosantes (Stylosanthes guianensis (Aubl.) Sw.). A tolerância a Al+3 em leguminosas forrageiras é relativamente menor, atingindo níveis semelhantes aos de cultivares comerciais de Medicago sativa (TESFAYE et al., 2001). Entretanto, a variabilidade presente no germoplasma para o caráter, sua natureza genética e os mecanismos envolvidos permanecem desconhecidos. A introduçăo de genes exógenos ou o aumento na expressăo de genes endógenos promove aumentos significativos na tolerância ŕ presença de altas concentraçőes de Al+3 solúvel no solo para diversas culturas (DE LA FUENTE et al., 1997; DELHAIZE ET AL., 2001; TESFAYE et al., 2001; DELHAIZE et al., 2004). O emprego destas técnicas pode avançar significativamente programas de melhoramento de espécies selvagens ou pouco melhoradas, disponibilizar mais rapidamente e com menor custo o material melhorado e introduzir maior variabilidade genética do caráter no germoplasma (QUECINI, 2006). Resistęncia ao frio e ŕ seca A resistęncia ao frio e ŕ seca visa introduzir genes de plantas resistentes em plantas de interesse comercial. Alguns exemplos podem ser: Em regiőes áridas, as plantas săo caracteristicamente espinhosas, mais resinosas ou mais tóxicas, uma vez que estăo expostas aos predadores em condiçőes de grande procura. Adaptaçőes morfológicas a diferentes climas săo conhecidas como aumento na quantidade de água armazenada nos tecidos ou grossas de cera para reduzir a perda de água. Adaptaçőes bioquímicas também săo conhecidas. Para reduzir a perda de umidade, as plantas fecham seus estômatos (aberturas na epiderme de folhas e caule, através das quais se efetuam as trocas gasosas necessárias ŕ vida das plantas). O fechamento ou abertura de estômatos obedece a um controle hormonal efetuado pelo ácido abscísico, uma substância sesquiterpenóide comum em plantas. Plantas tropicais, como a cana-de-açúcar, por exemplo, săo capazes de resistir ao clima quente e manter-se produtivas graças a uma modificaçăo no seu mecanismo fotossintético: o gás carbônico, que seria perdido por fotorrespiraçăo na superfície da folha, é coletado e trazido de volta ao interior do cloroplasto, onde se processa a fotossíntese, para ser convertido em sacarose. Plantas desse tipo tęm células mesofílicas especiais e suas modificaçőes anatômicas, conhecidas como Síndrome de Kranz, ocorrem paralelamente com as alteraçőes bioquímicas. http://www.geocities.com/~esabio/interacao/adapto_bioqu.htm Nas plantas superiores, adaptaçăo ao gelo parece ser bem mais complexa, mas sempre se faz uma correlaçăo da tolerância ao frio com o aumento do teor de açúcar na seiva. Provou-se que a capacidade de resistir ŕ geada pode ser induzida artificialmente, infiltrando-se plantas com açúcares. Os açúcares encontrados nas plantas resistentes ao congelamento variam de planta para planta, mas geralmente săo os açúcares mais conhecidos: glicose, frutose e sacarose. Álcoois polihídricos, como glicerina, manitol e sorbitol săo menos encontrados, mas săo largamente responsáveis pela tolerância ao frio da maçă e da gardęnia, por exemplo. Assim, conhecendo os mecanismo de resistęncia ao frio e seca, é possível garimpar genes para se fazer OGMs com estas características. Resistęncia a salinidade Um exemplo de condiçăo de solo adversa para o desenvolvimento de uma planta é a salinidade. Quando o teor de cloreto de sódio é aumentado, elas desenvolvem sintomas de intoxicaçăo, rapidamente. É o que acontece com as plantas chamadas "glicófitas", como o tomate, a ervilha e o feijăo. As plantas halófitas, pelo contrário, tęm seu crescimento estimulado em condiçőes de salinidade, como acontece com plantas costeiras tropicais, como as Thalassia e as plantas de mangue, como Avicennia (no Brasil, uma delas é a Rhizophora mangle).A resistęncia se dá por acúmulo de cloreto de sódio dentro do vacúolo; por resistęncia ŕ entrada de cloreto de sódio na célula e por diluiçăo de cloreto de sódio após sua entrada na planta. Uma característica bioquímica é o acúmulo de prolina e glicibetaína. http://www.geocities.com/~esabio/interacao/adapto_bioqu.htm O efeito do estresse salino pode provocar um estímulo ou inibiçăo de enzimas envolvidas nos processos metabólicos como a peroxidase, isoenzimas associadas a mudanças nos processos fisiológicos de plantas submetidas a estresses. Desde que as poliaminas e as peroxidases podem ser consideradas hipoteticamente como marcadores bioquímicos que mostram a resistęncia dos vegetais ŕ fatores abióticos, o objetivo do presente trabalho foi o de verificar alteraçőes nos níveis de poliaminas e na atividade da peroxidase em plantas de feijăo cultivadas em diferentes concentraçőes de NaCl. (Lima et al, 1999) Agradecimentos
Vera Quecini, PqC IAC pela cessăo de imagens
REFERĘNCIAS BIBLIGRÁFICAS CASTRO, A.M.G.; LIMA, S.M.V.; LOPES, M.A. et al. Estratégia de P & D para o melhoramento genético em uma época de turbulęncia. In: SIMPÓSIO DE GESTĂO DA INOVAÇĂO TECNOLÓGICA, 22. 2002. Salvador, Anais..., Salvador: PGT-USP/FIA/UNIFACB/FIEB, 2002. p. 1-17. CORNELISSEN, J. C. E MELCHERS, L. S. (1993) ‘Strategies for control of fungal diseases with transgenic plants’, Plant Physiol, 101:709-12. DALL’AGNOL, M.; SCHEFFER-BASSO, S.M. Utilizaçăo de recursos genéticos de leguminosas para ruminantes. In: REUNIĂO ANUAL DA SOCIEDADE BRASILEIRA DE ZOOTECNIA, 41. 2004. Campo Grande, Anais..., Campo Grande: SBZ/EMBRAPA Gado de corte, 2004. p. 115-129. DE LA FUENTE, J.M.; RAMÍREZ-RODRÍGUEZ, V. ; CABRERA-PONCE, J.L. ; HERRERA-ESTRELLA, L. Aluminun tolerance in transgenic plants by alteration of citrate synthesis. Science, v.276, p.1566-1568, 1997. DELHAIZE, E.; HEBB, D.M.; RYAN, P.R. Expression of a Pseudomonas aeruginosa citrate synthase gene in tobacco is not associated with either enhanced citrate accumulation or efflux. Plant Physiol., v.125, p.2059-67, 2001. DELHAIZE, E.; RYAN, P.R.; HEBB, D.M.; YAMAMOTO, Y.; SASAKI, T.; MATSUMOTO, H. Engineering high-level aluminum tolerance in barley with the ALMT1 gene. Proc. Natl. Acad. Sci. USA, v.101, p.15249-54, 2004. EVANS, D.A. Use of Protoplast Fusion for Crop Improvement. In: CROCOMO, O.J.; SHARP, W.R. & MELO, M. Biotecnologia para a Produçăo Vegetal. Piracicaba: CEBTEC/FEALQ, 1991. FINER, J.J.; FINER, K.R. & SANTARéM, E.R. Plant Cell Transformation, physical methods for. In: MEYERS, R.A. Encyclopedia of Molecular Biology and Molecular Medicine. Weinheim: VCH Publishers, 1996. FREYSSINET, G. E DEROSE, R. T. (Nov.-Dec. 1994) ‘Development of genetically modified crops resistant to herbicides and pests’. Agro-Food-Industry Hi-tech, 3-7. FROMM, M.E.; MORRISH, F.; ARMSTRONG, C. et al. Inheritance and Expression of Chimeric Genes in the Progeny of Transgenic Maize Plants. Bio/Technology, v.8, pp.883-839, 1990. GELVIN, S. B. (1998) ‘The introduction and expression of transgenes in plants’. Curr. Opin. Biotechnol., 9:227-32. GONSALVES, D. & SLIGHTOM, J. L. (1993) ‘Coat protein-mediated protection: analysis of transgenic plants for resistance in a variety of crops’. Seminars in Virology, 4:397-406. HADIDI, A.; KHETARPAL, R. K. E KOGANEZAWA, H. (1998) Plant virus disease control. APS Press, St. Paul, Minnesota, 677 p. KNOWLES, B. H. & DOW, J. A. T. (1993) ‘The crystal deltaendotoxinas of Bacillus thuringiensis: models for their mechanism of action on the insect gut’. Bio Essays, 15: 469-76. KOMARIT, T.; HIEI, Y.; SAITO, Y.; MURAI, N. E KUMASHIRO,T. (1996) ‘Vectors carrying two separate T-DNAs for co-transformation of higher plants mediated by Agrobacterium tumefaciens and segregation of transformants free from selection markers’. Plant. J. 10:165-74. LIMA, G.P.P.; BRAZĂO, O.G.; OLIVEIRA, A.M. Poliaminas e atividade da peroxidase em feijăo (phaseolus vulgaris l.) cultivado sob estresse salino. Sci. agric. v.56 n.1, 1999. LIPP-NISSINEN, K. Molecular and Cellular Mechanisms of Agrobacterium-mediated Plant Transformation. Cięncia e Cultura, Săo Paulo, 45: 104-111, 1993. LUTHRA, R. et al. Microprojectile mediated Transformation: a bibliographic search. Euphytica, Wageningen, 95: 269-294,1997. MCCABE, D.E. & MARTINELLI, B.J. Transformation of Elite Cotton Cultivars by Particle Bombardment of Meristems. Bio/Technology, Nova York, v.11, pp. 596-598, 1993. PEREIRA, A.V. Avanços no melhoramento genético de gramíneas forrageiras tropicais. In: REUNIĂO ANUAL DA SOCIEDADE BRASILEIRA DE ZOOTECNIA, 39. 2002. Recife, Anais..., Recife: SBZ/UFRPE, 2002. p. 19-41. PEREIRA, A.V. Melhoramento genético de plantas forrageiras. In: SIMPÓSIO SOBRE ATUALIZAÇĂO EM GENÉTICA E MELHORAMENTO DE PLANTAS, 2. 1998. Lavras, Anais..., Lavras: UFLA/FAEP, 1998. p. 135-162. PEREIRA, A.V.; VALLE, C.B.; FERREIRA, R.P. et al. Melhoramento de Forrageiras Tropicais. In: NASS, L.L.; VALOIS, A. C.C.; MELO, I.S.; et al (Ed.). Recursos genéticos e melhoramento de plantas: Rondonópolis: Fundaçăo MT, 2001. p. 549-602. PINHEIRO, M. M.; GERHARDT, L.; MARGIS, R.: ‘Uma tecnologia com múltiplas aplicaçőes’. História, Cięncias, Saúde — Manguinhos, vol. VII(2), 465-79, jul. out. 2000. PINTO, F.S.A. Hibridaçăo em soja. www.ufv.br/dbg/bioano02/a2001a05.htm, 2001. QUECINI, V. Introduçăo de Tolerância a Alumínio em forrageiras: soja perene (Neonotonia wightii (Wight & Arn.) Lackey), leucena (Leucaena leucocephala (Lam.) de Wit) e braquiária (Brachiaria brizantha (C. Hochstetter ex A. Rich.) Stapf.). Relatório de Atividades, março de 2006. Doc. IZ/MATSUDA. SANTARÉM, E.R. & FERREIRA, A.G. Transformaçăo de Soja via Bombardeamento de Partículas. ABCTP Notícias, Brasília, 9: 2-9, 1997. SANTARéM, E.R. et al. Sonication-Assisted Agrobacteriummediated Transformation of Soybean Immature Cotyledons: optimization of transient expression. Plant Cell Reports, Berlin, 17: 752-759. 1998. SANTARÉM, E.R. Métodos eficientes para transformaçăo de plantas. www.unimep.br/phpg/editora/revistaspdf/rct15art10.pdf+bo mbardeador+de+part%C3%ADculas&hl=pt-BR&gl=br&ct=clnk&cd=1. 2003. SHADE, R. E.; SCHROEDER, H. E.; PUEYO, J. J.; TABE, L. M.; MURDOCK, L. L.; HIGGINS, T. J. V. E CHISPEELS, M. J. (1994) ‘Transgenic pea seeds expressing the (-amilase inhibitor of the commonbean are resistant to bruchid beetles’. Bio/technology, 12:796-800. SHELTON, H.M.; FRANZEL, S. & PETERS, M. Adoptions of tropical legume technology around the world: analysis of success, 149-166 in: Grassland A Global Resource ed. McGilloway, Irlanda, 2005. SOUTHGATE, E.M. et al. A Comparison of Methods for Direct Gene Transfer into Maize (Zea mays L.). In Vitro Cellular and Developmental Biology-Plant, Mariland, 34: 218-224, 1998. TESFAYE, M.; TEMPLE, S.J.; ALLAN, D.L.; VANCE, C.P.; SAMAC, D.A. Overexpression of malate dehydrogenase in transgenic alfalfa enhances organic acid synthesis and confers tolerance to aluminum. Plant Physiol., v.127, p.1836-44, 2001. VASIL, V.; LU, C.Y. & VASIL, I.K. Histology of Somatic Embryogenesis in Cultured Immature Embryos of Maize. Protoplasma, Wien, 127: 1-8, 1985. VIEIRA, L.G.E.OGMs -uma tecnologia controversa. Cięncia Hoje, abril 2004. www.cienciahoje.com.br . WOOD, S.; SEBASTIAN, K.; SCHERR, S. Soil resource condition. In Pilot Analysis of Global Ecosystems: Agroecosystems. International Food Policy Research Institute and the World Resources Institute, Washington, DC, pp.45-54, 2000. YAMAME, R.S. Marcadores Moleculares. www.ufv.br/dbg/trab2002/ GMOL/ GMOL 003.htm, 2002. ZAMBRYSKI, P. Basic Processes Underlying Agrobacteriummediated DNA Transfer to Plant Cells. Annual Review of Genetics, Cambridge, 22: 1-30, 1988.
Sites Consultados: http://cienciahoje.uol.com.br http://www.geocities.com/~esabio/interacao/adapto_bioqu.htm
Paulo
Bardauil Alcântara
possui graduaçăo em Agronomia pela Universidade de Săo Paulo (1974),
especializaçăo em COLHEITA DE FORRAGEM E UTILIZAÇĂO DE PASTAGENS pela
Volcani Center (1976), mestrado em Zootecnia pela Universidade Estadual
Paulista Júlio de Mesquita Filho (1981) e pós-doutorado pela University
of Queensland (1990). Atualmente é DIRETOR de Pesquisa /PQC VI do
Instituto de Zootecnia da APTA/SAA. Tem experięncia na área de Zootecnia
, com ęnfase em Pastagem e Forragicultura.
Keila Maria Roncato Duarte
possui graduaçăo em Engenharia Agronômica pela Universidade de Săo Paulo
(1992), mestrado em Microbiologia Agrícola pela Universidade de Săo
Paulo (1995) e doutorado em Microbiologia Agrícola pela Universidade de
Săo Paulo (2000). Atualmente é Pesquisadora Científica da APTA -
Instituto de Zotecnia. Fez pós-doutorado no CENA/USP na área de produçăo
de anticorpos policlonais e monoclonais para detecçăo de anabolizantes
em carnes (2000 A 2003). De 2003 A 2005, Fez um segundo pós doutorado na
USP, no Departamento de Fitopatologia, Entomologia e Zoologia Agrícola -ESALQ
na área de epidemiologia e imunomarcaçăo de doenças fúngicas. Tem
experięncia na área de Imunologia aplicada e microbiologia, com ęnfase
em Produçăo de Anticorpos Monoclonais e Policlonais, atuando
principalmente nos seguintes temas: anticorpos policlonais, anticorpos
monoclonais, anabolizantes , Imunomarcaçăo, diagnóstico de fitopatógenos,
ELISA. Hoje atua no Laboratório de Reproduçăo Animal, com imunoensaios
aplicados a estudos de fisiologia bovina, principalmente.
Waldssimiler Teixeira de Mattos possui
graduaçăo em Engenharia Agronômica pela Universidade de Săo Paulo
(1993), mestrado em Solos e Nutriçăo de Plantas pela Universidade de Săo
Paulo (1997), doutorado em Solos e Nutriçăo de Plantas pela Universidade
de Săo Paulo (2001) e pós-doutorado pelo Instituto de Zootecnia (2004).
Atualmente é Pesquisador Científico do Instituto de Zootecnia. Tem
experięncia na área de Zootecnia , com ęnfase em Pastagem e
Forragicultura. Atuando principalmente nos seguintes temas: forrageira,
adubaçăo.
Dados para citaçăo bibliográfica(ABNT): ALCÂNTARA, P.B.; DUARTE, K.M.R; MATTOS, W.T. Novas alternativas para germoplasma forrageiro - leguminosas. 2007. Artigo em Hypertexto. Disponível em: <http://www.infobibos.com/Artigos/2007_2/leguminosas/index.htm>. Acesso em: Publicado no Infobibos em 25/06/2007
|