|
A MATURAÇĂO FISIOLÓGICA DA VIDEIRA VINÍFERA EM SĂO PAULO: Comentários sobre as safras de verăo e de inverno na média altitude paulista
Antonio Odair Santos A qualidade do vinho é diretamente ligada ao ponto ótimo da maturaçăo da uva, sendo este um evento que envolve a maturaçăo fisiológica (biossíntese evolucionária na baga), da maturaçăo tecnológica (acúmulo de açúcar + ácidos) e da maturaçăo fenólica (acúmulo quali-quantitativo de taninos, pigmentos e compostos ligados ao sabor e aroma) (Mandelli et al. 2003). A localizaçăo do vinhedo e o seu manejo tęm fundamental importância neste aspecto. A investigaçăo sobre o tema tem sido feita para identificar estratégias de manejo que possam levar a otimizaçăo da maturaçăo, contudo para o melhor entendimento sobre estas estratégias faz-se necessário considerarmos a formaçăo e desenvolvimento da baga. Para isso é necessário observar primeiro as condiçőes críticas da sua formaçăo e as influęncias de variáveis microclimáticas sobre o processo de maturaçăo fisiológica.
O fruto da videira é essencialmente uma fábrica bioquímica. Além da síntese dos metabólitos essenciais primários (água, aminoácidos, minerais, micronutrientes) a baga tem a habilidade de sintetizar outros componentes que podem definir um vinho em particular (p.e. componentes de sabor e aroma) (Gholami et al 1995).
Há um potencial para uma grande variabilidade na maturaçăo da produçăo da videira, dentro do cacho, de cacho a cacho e dentro do vinhedo. Na prática é difícil determinar quando um vinhedo um pouco mais extenso está expressando o melhor potencial de maturaçăo. Um dos objetivos, no velho e novo mundo, tem sido produzir uma vindima o mais uniforme possível, quanto aos índices de maturaçăo (Wample et al, 2006; Santos et al, 2008). Neste espírito a melhor vindima seria aquela que resulta de um vinhedo com maturaçăo uniforme, bagas pequenas com a maturaçăo tecnológica da baga “par e passo” com a maturaçăo fenólica.
A baga é alimentada através do pedicelo por um sistema vascular composto do xilema e floema. O xilema é o sistema de vasos responsável pelo transporte da água, minerais, reguladores do crescimento e nutrientes do sistema radicular para o resto da planta. Evidęncias atuais indicam que o xilema é funcional na baga até a “pinta”; posteriormente esta funçăo é reduzida ou eliminada. A baga também é suprida pelo floema, que é o sistema vascular transportador dos fotossintetizados (sucrose) das folhas para a planta; esta funçăo é reduzida no início do desenvolvimento da baga, mas tornase a principal fonte de ingresso após a “pinta”(Greenspan et al 1994).
O aumento no volume da baga, primariamente devido ŕ água, está associado ao aumento de açúcar após a “pinta”. Contudo em algumas variedades (notadamente Syrah), principalmente se cultivada sob clima quente e seco, o aumento em açúcar durante os estágios finais da maturaçăo năo é acompanhado pelo aumento do volume da baga, mas é causado pelo murchamento da mesma. Esse murchamento parece estar relacionado com a transpiraçăo da baga (McCarthy, 1999; Santos & Kaye, 2009), o que sugere que a inabilidade da baga para permanecer em ótima hidrataçăo nesse ponto do ciclo é devido ao bloqueio de elementos do floema dentro da baga. Suspeita-se que, para algumas variedades, o sistema vascular entre a videira e a baga tem a funçăo reduzida durante a parte final da maturaçăo da baga.
Desenvolvimento das fases inicias da baga O desenvolvimento da baga consiste de duas curvas sigmóides sucessivas de crescimento, separadas por um “retardo”. O primeiro período de crescimento tem início na floraçăo, estendendo-se até aproximadamente 60 dias. Durante o primeiro período de crescimento a baga é formada e os embriőes da semente săo produzidos; uma rápida divisăo celular ocorre nas primeiras semanas e, no final deste período, o total do número de células da baga está estabelecido (Harris et al., 1968). A dimensăo na divisăo celular tem importância no eventual tamanho da baga e, a dinâmica da água no solo influencia esse processo. Há vários solutos que acumulam na baga durante este primeiro período, e todos aparentemente atingem um máximo ao redor da “pinta” (Possner et al.,1985). Os mais importantes săo o ácido málico e tartárico, sendo que o ácido tartárico é o que é sintetizado primeiro, no inicio do período. Estes ácidos compőem a acidez do vinho e, portanto, săo críticos para a qualidade do mesmo. Igualmente, no primeiro período acumulam-se os ácidos hidroxicinámicos, importantes, devido ao seu envolvimento com reaçőes de escurecimento e porque săo precursores de fenóis voláteis (Licker et al., 1999). Os taninos, incluindo as catequinas monoméricas também se acumulam no primeiro período de crescimento da baga. Estăo presentes na casca e na semente e quase ausentes na polpa; săo responsáveis por caracteres de amargor e adstringęncia no vinho e também importantes na fixaçăo da cor (Kennedy et al., 2001). Há outros componentes, năo menos importantes para a qualidade do vinho, que se acumulam na primeira fase de crescimento: minerais, aminoácidos, micronutrientes e componentes do aroma (p.e. metoxipirazinas).
Desenvolvimento do segundo período de crescimento O início da segunda fase de crescimento da baga coincide com a “pinta” e é caracterizado pelo amolecimento e coloraçăo do fruto; praticamente dobra em tamanho entre o início da segunda fase de crescimento e a colheita. Muitos dos solutos que acumularam na primeira fase de crescimento permanecem na colheita; contudo, devido ao aumento de volume da baga, a concentraçăo é reduzida significantemente. Alguns compostos produzidos durante o primeiro estágio de crescimento da baga săo reduzidos (e năo diluídos) durante a segunda fase decrescimento (g/baga). Entre eles, o ácido málico, usado como fonte de energia durante o processo de maturaçăo. Esta reduçăo varia consideravelmente, mas pode ser empiricamente correlacionado com as condiçőes climáticas. Videiras cultivadas em regiőes quentes tendem a ter menos ácido málico que aquelas cultivadas em regiőes frias; taninos também declinam consideravelmente durante a segunda fase de crescimento (g/baga). Os taninos presentes na semente aparentemente sofrem oxidaçăo ao tempo em que se tornam fixos na pele da semente. Como resultado, a composiçăo dos taninos extraíveis das sementes mudam consideravelmente e é caracterizada pela reduçăo proporcional nos componentes tânicos amargos (Kennedy et al., 2000). Os taninos da casca declinam ou permanecem constantes durante a segunda fase de crescimento da baga e igualmente sofrem modificaçőes, sobretudo pela açăo da antocinanina e pectinas, o que tem implicância para a textura e estabilidade da cor no vinho.
Notadamente, componentes aromáticos que săo sintetizados na primeira fase de crescimento da baga declinam (g/baga) durante o amadurecimento da baga. Isto inclui vários dos compostos de metoxipirazinas, responsáveis pelo caráter vegetal/herbáceo de alguns vinhos (p.e. Cabernet Sauvignon e Sauvignon Blanc). O declínio nos níveis de pirazinas está relacionado com os níveis de radiaçăo solar incidente na zona de frutificaçăo e com a dinâmica da água no solo, dentre outros fatores (Hashizume e Samuta, 1999). Portanto, ocasionalmente sendo estes compostos indesejáveis em certos níveis, o manejo no vinhedo pode ser usado para reduzi-los.
Na segunda fase de crescimento tem-se um grande aumento em compostos (a maior parte glicose e frutose) que ocorre como resultado da mudança bioquímica no modo de maturaçăo do fruto. No começo da “pinta” o fluxo de açúcar para a baga inicia, sendo que a sucrose produzida na fotossíntese é importada para dentro da baga e hidrolizada para glicose e frutose (Robinson & Davis, 2000). A eventual concentraçăo também depende de outros fatores como variedade, regime microclimático, índice de carga, dimensăo do dossel, estado fitossanitário e eventual sobrematuraçăo.
Em que pese o acúmulo de açúcar ser importante, o maior determinante da qualidade do vinho săo os metabólitos secundários. Nas variedades tintas a produçăo de antocianina, restrita a casca do fruto na maioria das cultivares, é o composto de importância mais obvia; doravante, tanto quanto para variedades brancas a maioria dos compostos aromáticos voláteis săo produzidos durante esta fase da maturaçăo do fruto (Wilson et al., 1984). Estes últimos se referem aos compostos da classe dos terpenóides (a maior parte monoterpenos) que săo importantes aromas florais presentes em muitas variedades, como Riesling e ‘Moscatos’, juntamente com precursores de aromas frutados. Os componentes do aroma săo distribuídos na polpa e na casca do fruto; alguns destes săo apenas precursores, estando presentes na baga como glicosidios, năo sendo voláteis (quando săo) até que o vinho tenha sido produzido e submetido a guarda (Francis et al., 1999).
Impactos do
microclima sobre maturaçăo fisiológica e fenólica A maturaçăo da baga é altamente dependente da temperatura do ar, assim como qualquer reaçăo bioquímica-enzimática. O processo de maturaçăo do fruto tem a temperatura ótima, abaixo ou acima do qual o processo perde intensidade. A curva de resposta a temperatura para cada um dos processos descritos năo é totalmente entendida. Contudo, ela ocorre, provavelmente, ao redor de 30 a 32 şC. Acima destes limites as taxas decrescem rapidamente. Abaixo destes valores as taxas decrescem com menor aceleraçăo, cessando ao redor de 10 şC.
A soma térmica na estaçăo de crescimento pode ser verificada através do conceito de graus-dia (Pedro-Jr et al., 2004). Desta maneira pode se monitorar a atividade biológica do fruto durante a estaçăo de crescimento. A soma térmica é útil como indicador da capacidade do local para a maturaçăo de determinada variedade. Contudo, é importante observar o padrăo comportamental da soma térmica . É possível que duas localidades com a mesma soma térmica tenham diferentes janelas climáticofenológicas. Particularmente, latitudes maiores ou regiőes de altitude frequentemente tęm baixa soma térmica em ambas a pontas (fim e inicio da estaçăo) fazendo-se necessário usar variedades que possam tolerar uma janela climática mais curta.
A soma térmica é um bom indicador inicial da influęncia climática sobre um vinhedo em particular. Mas, pode năo ser suficiente para descrever como o microclima influencia a maturaçăo, especificamente. A média diurna da temperatura do ar durante a maturaçăo é útil para este propósito. A diferença entre a temperatura do ar diurna e noturna, assim como as temperaturas míninas nos dois períodos săo reveladoras das condiçőes para a maturaçăo de um vinhedo em um local particular.
O velho ditado “dias quentes e noites frias” năo necessariamente descreve uma situaçăo ideal. Contudo se uma regiăo tem dias quentes, noites frias săo essenciais para prevenir o rápido metabolismo ácido. Um clima com dias quentes e noites quentes, produz um diferente (năo necessariamente inferior) estilo de vinho, na comparaçăo com uma regiăo ainda com a mesma soma térmica, mas desta feita, com dias quentes e noites frias. Quando temperaturas noturnas se aproximam de 10 graus Celsius a baga cessa a maturaçăo. Quando a temperatura noturna eleva-se acima daquele patamar, a maturaçăo continua dentro da noite. Assumindo-se que a temperatura diurna aproxima-se do ótimo para maturaçăo, a menor flutuaçăo térmica diária produz a maturaçăo mais rápido que climas com larga flutuaçăo térmica diária. Na prática, para uma determinada variedade, microclimas com menor flutuaçăo térmica diária geralmente matura o fruto a taxas mais altas de sucrose, na comparaçăo com climas com larga flutuaçăo térmica e maiores índices de nebulosidade e com similar soma térmica.
Por outro lado, a radiaçăo solar tem grande importância no desenvolvimento de antocianina na casca do fruto das variedades tintas. Por isso um dossel congestionado ou uma zona de frutificaçăo muito coberta tende a produzir coloraçăo pobre na baga. Contudo, somente cerca de 10 % do total da radiaçăo solar global incidente é necessário para produzir o efeito desejado (Deloire & Hunter, 2005). Aumentando-se os níveis de radiaçăo solar pode-se incorrer em riscos de escaldadura.
A luz ultravioleta também tem mostrado influenciar a concentraçăo de flavonóis na baga. Esse efeito foi provado ser separado do efeito da temperatura (Spayd et al., 2002). Os flavonóis tais como quercetina e kaempferol săo protetores solares naturais da baga; săo importantes antioxidantes com importante contribuiçăo medicinal no vinho; a contribuiçăo em sabor e aroma no vinho năo é garantida, mas podem em certos casos influenciar a percepçăo sensorial de outros elementos aromáticos.
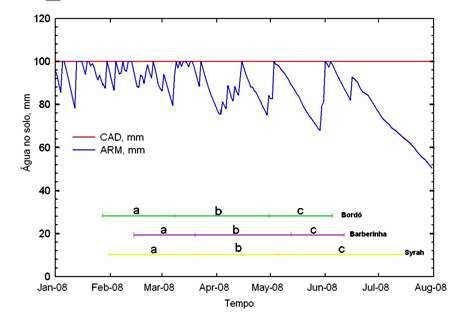
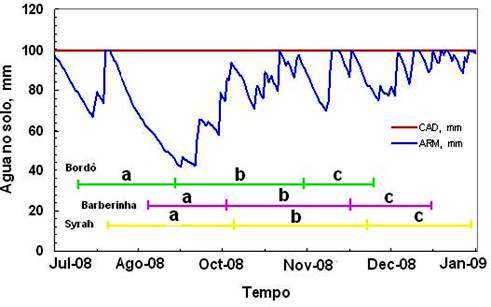
As Tabelas 1 e 2 se referem aos dados do ciclo da videira colhida no verăo (poda normal, no inverno) e colhida no inverno (poda no verăo) em Săo Paulo. Os dados dăo uma boa idéia sobre as condiçőes de maturaçăo da uva na média altitude paulista. Verifica-se que a maturaçăo fisiológica da baga é jogada em quadrantes diferentes das estaçőes do crescimento, no que se refere a soma térmica e disponibilidade hídrica, o que resulta em condiçőes diferenciadas para a maturaçăo tecnológica e fenólica da uva.
Para a safra de verăo (SV) a soma térmica disponível para cumprir o ciclo é maior que a safra de inverno (SI), sendo que a flutuaçăo térmica diurna é menor nesta última, o que contribui para que ‘Syrah’ tenha o ciclo expandido. Por outro lado, a intensidade de cor é maior na colheita de inverno, sendo que a formaçăo e maturaçăo da baga ocorreram sob condiçőes de disponibilidade hídrica declinante (Figura 1); em comparaçăo com o cultivo do verăo, ocorre o contrário, verificando-se o armazenamento hídrico crescente, no período de maturaçăo (Figura 2). Notadamente, se tem para a safra de verăo e de inverno um número de dias com chuva semelhantes nos dois eventos, porém a concentraçăo da chuva se dá em épocas diferentes do ciclo, o que faz a diferença. Este detalhe tem particular importância e mostra que as condiçőes para a maturaçăo fisiológica e fenólica da videira no outono/inverno (SI) săo mais favoráveis, principalmente para Vitis vinifera. Para as variedades americanas e híbridas, menos exigentes que as viníferas verdadeiras, as condiçőes da SV e da SI tendem produzir resultados menos contrastantes, considerando-se somente a disponibilidade térmica e hídrica; a pressăo fitopatológica esperada também é menor na SI, muito embora o desenvolvimento dos ramos da videira, neste período, ainda coincida com um período de precipitaçăo mais intenso.
Os dados sobre peso dos cachos e o peso de bagas evidenciam que para SI a produçăo é menor, com tamanho de baga reduzido. Provavelmente a disponibilidade hídrica na primeira fase de crescimento da mesma é fator determinante para o decréscimo da dimensăo da baga, devido a que nesta fase está localizada a maior intensidade da divisăo celular, em relaçăo ŕ expansăo, conforme discutido acima. De fato, em relaçăo ŕ Safra de verăo o cultivo de outono/inverno tem o estabelecimento do fruto e as primeiras fases da formaçăo da baga coincidindo com o final das chuvas de verăo; exatamente o contrário ocorre no período da SV, conforme se constata pelos valores de armazenamento da água no solo (Figuras 1 e 2).
Os dados apresentados sugerem que para alguns casos pode ser adequada a recomendaçăo da inversăo de safra para Vitis vinifera, com a derrubada de cachos na formaçăo dos mesmos e o cultivo em dupla poda seqüencial, com aproveitamento das duas safras, para as cultivares “americanas” e “híbridas”, na média altitude paulista. Muito embora, mais dados ainda devam ser acrescentados a esta linha de investigaçăo no futuro, as Tabelas 1 e 2, em conjunto com as Figuras 1 e 2 sugerem melhores condiçőes para maturaçăo tecnológica e maturaçăo fenólica da uva na safra de inverno, principalmente para os anos “típicos” onde o cultivo de outono/inverno ocorre sem as flutuaçőes térmicas causadas pela dinâmica de massa polares, que em alguns anos podem estacionar temporariamente sobre a regiăo.
Por outro lado, a inversăo de safra citada requer a determinaçăo do índice de carga mais favorável ŕ otimizaçăo da maturaçăo e um cuidado bastante rigoroso na adubaçăo do vinhedo. Além disso, um rodízio de áreas de produçăo é recomendado, para năo sobrecarregar o vinhedo, o que poderia causar a queda na vida útil do mesmo. O controle do vigor pode ser feito com o manejo do dossel e/ou manejo no desbaste de cachos, buscando-se o melhor ajuste da carga. A velha máxima de que vigor é qualidade da maturaçăo săo mecanismos inversos pode ser verdade, mas para o microclima da média altitude paulista faz-se necessário primeiro estabilizar o vinhedo e depois controlar o índice de carga e o vigor vegetativo. Năo obstante, os solos mais pobres poderăo render os melhores vinhos, justamente porque aí se tem a chance de melhor controlar a disponibilidade de nutrientes e água. Conforme foi discutida a maturaçăo tecnológica e fenológica da videira ocorre dentro da baga, sob processos altamente influenciados pelo meio físico, o que tem ainda mais importância quando tratamos de Vitis vinifera.
Na média altitude paulista, existem condiçőes críticas que demandam rigor na conduçăo da videira, para melhor aproveitamento das condiçőes do meio objetivando a alcançar melhores índices de maturaçăo; cita-se o vento, o quadrante do ano de maior disponibilidade hídrica, as janelas térmicas de verăo e outono/inverno e os solos da regiăo que, em alguns casos, podem armazenar água mais tempo do que o desejado para a melhor qualidade da uva e do vinho local.
Agradecimento Vinícola Família Miqueletto – Louveira (SP) Vinícola Família Lourençon – Itupeva (SP)
Referęncias
DELOIRE, A.; HUNTER, J.J. Microclimat
des grappes et maturation du raisin berry composition as affected by
bunch exposure. Progrčs Agricole et Viticole, v 122, n7, pg.
200-206, 2005.
DELOIRE, A.; OJEDA, H.; ZEBIC, O.;
BERNARD, N.; HUNTER, J.J.; CARBONNEAU, A. Influence de l´état
hydrique de la vigne sur le style de vin. Progrčs Agricole et
Viticole, v. 122, n.21, p. 455-462, 2005.
FRANCIS, I.L.; KASSARA, S.; NOBLE,
A.C.; WILLIAMS, P.J. The contribution of glycoside precursors to
Cabernet Sauvignon and Merlot aroma. ACS, Washington, 1999. p.13-30.
GHOLAMI, M.; HAYASAKA, Y.; COMBE,
B.G.; JACKSON, J.F.; ROBINSON, S.P.; WILLIAMS, P.J. Biosynthesis of
flavour compounds un Muscat Gordo Blanco grape berry. Australian
Journal of Grape & Wine Research, v.1, p. 19-24, 1995.
GREENSPAN, M.D.; SHACKEL, K.A.;
MATTHEWS, M.A. Development changes in the diurnal water budget of
the grape berry exposed to water deficits. Plant Cell & Environment,
v. 17, p. 811-820, 1994.
HARRIS, J.M.; KRIEDMANN, P.E.;
POSSINGHAM, J.V. Anatomical aspects of grape berry development.
Vitis, n.7, p.106-109,1968.
HASHIZUME, K.; SAMUTA, T. Grape
maturity and light exposure affect berry methoxypyrazine
concentration quality. American Journal of Enology and Viticulture,
v. 50,
p.194-198, 1999.
KENNEDY, J.A.; HAYASAKA, Y.; VIDAL S.;
WATER, E.J.; JONES, G.P. Composition of grape skin proanthocyanidins
at different stages of berry development. Journal of Agricultural &
Food Chemestry, v. 49, p. 5348-5355, 2001.
KENNEDY, J.A.; MATTHEWS, M.A.;
WATERHOUSE, A.L. Changes in grape seed polyphenols during fruit
ripening. Phytochemesty, v. 55, p.77-85,2000.
LICKER, J.L.; ACREE, T.E.;
HENICK-KLING, T. What is Brett (Brettanomyces) flavor? A preliminary
investigation. In: Chimestry of wine flavor. WATERHOUSE, A.L.;
EBELER, S.E. (Eds). ACS, Washington, 1999. p. 96-115.
MANDELLI, F. ; BERLATTO, M. A. ;
TONIETTO, J. ; BERGAMASCHI, H. . Fenologia da videira na Serra
Gaúcha. Pesquisa Agropecuária Gaúcha, Porto Alegre, v. 9, n. 1-2, p. 129-144, 2003.
McCARTHY, M.G. Weight loss from ripening grapevines (vitis
vinifera L. cv syrah).
Australian Journal of Grape & Wine
Research, v. 5, n.1, p.10-16, 1999. PEDRO JR., J.J.; CAMARGO, M.B.P.;
MORAES, A.V.C; FELÍCIO, J.C.; CASTRO,
J.L.
Temperatura base, graus-dia e duraçăo do ciclo para cultivares de
triticale. Bragantia, Campinas, v 63, n.3, p.447-453, 2004. POSSNER,
D.R.E.; KLIEWER, W.M. The localization of acids, sugars, potassium
and calcium in developing grape berries. Vitis, v.24, p. 229, 240.
ROBINSON, S.P.; DAVIES, C. Molecular biology of grape berry ripening.
Australian Journal of Grape and Wine Research, v. 6, n.2, p.
168.174, 2000.
SANTOS, A.O. ; KAYE, O.
Composiçăo quali-quantitativa da produçăo de syrah cultivada sob
estresse hidrico transiente. Revista Brasileira de Engenharia
Agrícola e Ambiental, v. 13, p. 272-281, 2009.
SANTOS, A.O.;WAMPLE, R. L. ;
SANDHANDAN, S. ; SETHURAMASAMYRAJA,
B. ;
KAYE, O. . First attempts at instrumenting for grape quality-based
harvesting procedure. In: CIGR International Conference of
Agricultural Engineering, 2008, Foz do Iguacú. XXVII CIGR
International Conference of Agricultural Engineering. Jaboticabal
-SP : SBEA, 2008. v. 1. p. CD ROM.
SPAYD, S.E.; TARARA, J.M.; MEE, D.L.;
FERGUSON, J.C. Separation of sunlight and temperature effects on the
composition of vitis vinifera cv merlot berries. American Journal of
Enology and Viticulture, n. 53, p. 171-182, 2002
WAMPLE, R. L.; SANTOS, A.O. ; KAYE, O.
. Implementation of calibrated near-infrared spectroscopy in
precision viticulture and selective harvesting of wine grapes.
American Journal of Enology and Viticulture, Sacramento, CA, p. 382a
– 395a, 10 dez. 2006. WILSON, B.; STRAUSS, C.R.; WILLIAMS, P.J. Changes in free and glycosidically bound monoterpenes in developing Muscat grapes. Journal of Agricultural & Food Chimestry, v. 32, p. 919-924, 1984. Origem: Instituto Agronômico - IAC - www.iac.sp.gov.br
Antonio Odair Santos
possui graduaçăo em Engenharia
Agronômica pela Universidade do Estado de Santa Catarina (UDESC);
Mestrado e Doutorado em Fitotecnia, pela Universidade Federal do Rio
Grande do Sul (UFRGS); Pós-Doutorado pela California State
University-Fresno (USA) [Viticulture and Enology Research Center].
Atualmente é Pesquisador do Instituto Agronômico (IAC), atuando
principalmente nos seguintes temas: agrometeorologia da videira,
manejo da videira e qualidade de uva/vinho Contato: odairsan@iac.sp.gov.br
Glauco de Souza Rolim
possui doutorado em Agronomia
(Irrigaçăo e Drenagem) pela Universidade Estadual Paulista Júlio de
Mesquita Filho (2003), mestrado em Agronomia (Física do Ambiente
Agrícola) pela Universidade de Săo Paulo (2000) e graduaçăo em
Engenharia Agronômica pela Universidade de Săo Paulo (1996).
Atualmente é pesquisador científico do Instituto Agronômico de
Campinas-IAC. Tem experięncia na área de Agronomia, com ęnfase em
Agrometeorologia, atuando principalmente nos seguintes temas:
Modelagem Agrometeorológica, Micrometeorologia, Balanço Hídrico,
Balanço de Energia, Risco Climático, Citros, Café e videira. Contato: rolim@iac.sp.gov.br
Jose Luiz Hernandes
é Técnico em Agropecuária, formado
pela ETAESG Benedicto Storani de Jundiaí (1987), graduado em
Cięncias com Habilitaçăo Plena em Biologia pela Faculdade de
Cięncias e Letras Padre Anchieta (1998) e mestrado em Agricultura
Tropical e Subtropical pelo Instituto Agronômico de Campinas (2001).
Em 2006 realizou o Curso Superior de Especializaçăo em Viticultura e
Enologia em Climas Quentes pelo Instituto de Formaçăo Agrária e
Pesqueira (CONSELHO DE INOVAÇĂO, CIĘNCIA E EMPRESA) da Junta de
Andaluzia, no Centro de Pesquisa e Formaçăo Agrária RANCHO DE LA
MERCED em Jerez de la Fronteira, Espanha. Atualmente é Pesquisador
Científico do Instituto Agronômico de Campinas. Tem experięncia na
área de Fitotecnia, com ęnfase em Vitivinicultura, atuando
principalmente nos seguintes temas: microclima, fitossanidade,
propagaçăo, sistemas de conduçăo, cultivo protegido, variedades de
videira para mesa e indústria e vinificaçăo artesanal. Contato: jlhernandes@iac.sp.gov.br
Mário José Pedro Júnior
possui
graduaçăo em Engenharia Agronômica pela Universidade de Săo Paulo
(1972), mestrado em Solos e Nutriçăo de Plantas pela Universidade de
Săo Paulo (1977) e doutorado em Agrometeorology - University Of
Guelph (1980). Atualmente é pesquisador científico do Instituto
Agronômico de Campinas, revisor de cinco revistas especializadas.
Tem experięncia na área de Agronomia, com ęnfase em
Agrometeorologia, atuando na área de fruticultura, principalmente
videira, nos seguintes temas: cultivo protegido, sistema de
conduçăo, microclima e zoneamento agrícola. Contato: mpedro@iac.sp.gov.br
Dados para citaçăo bibliográfica(ABNT): SANTOS, A.O.; ROLIM, G.S.; HERNANDES, J.L.; PEDRO JÚNIOR, M.J. A maturaçăo fisiológica da videira vinífera em Săo Paulo: comentários sobre as safras de verăo e de inverno na média altitude paulista. 2009. Artigo em Hypertexto. Disponível em: <http://www.infobibos.com/Artigos/2009_3/maturacao/index.htm>. Acesso em:Publicado no Infobibos em21/09/2009
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||